Trono Lab – Laboratory of Virology and Genetics
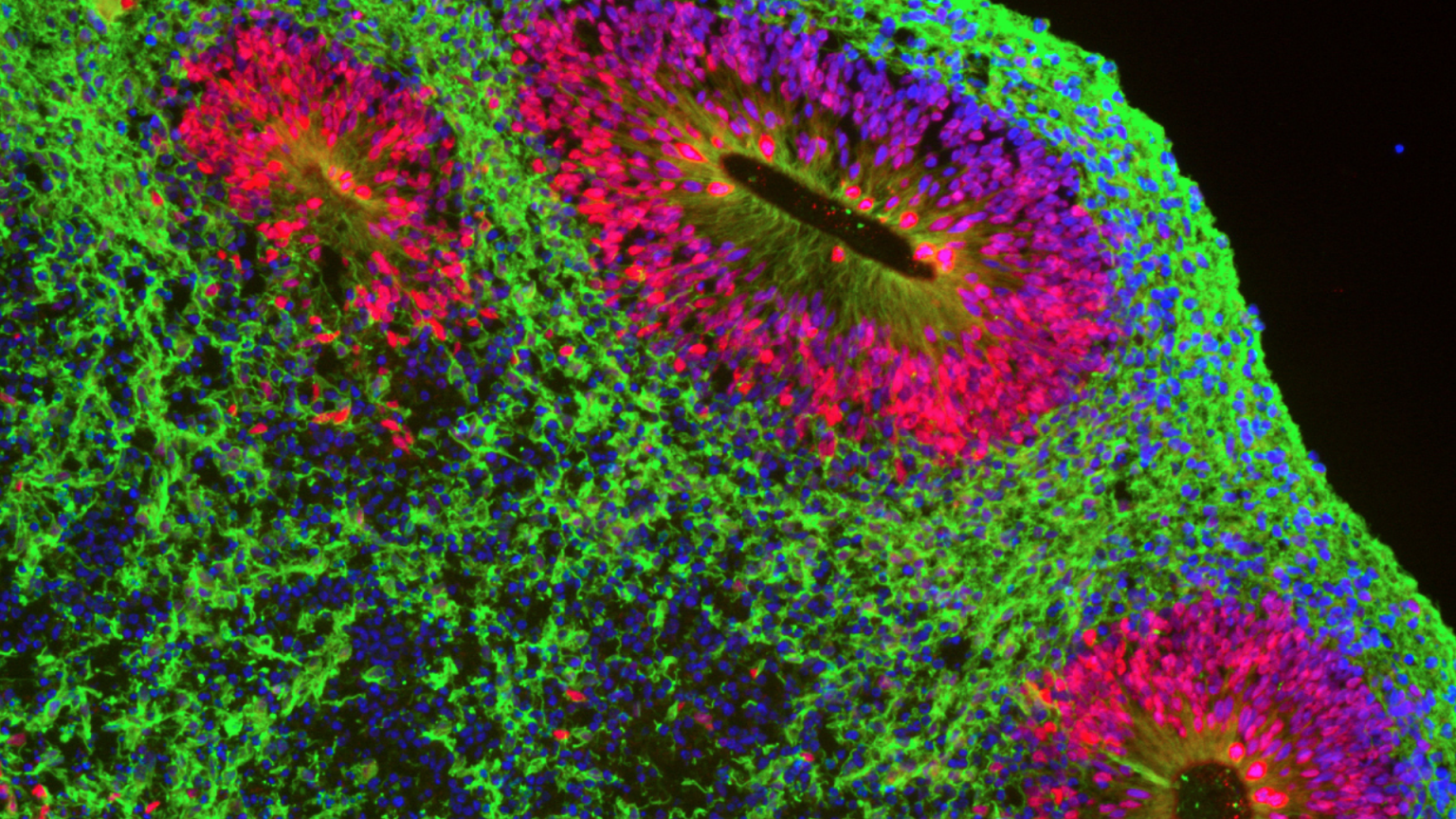
Our research interest are Krab-Zinc finger protein, transposons and epigenetics.

Contact
Head of Laboratory : Didier Trono
Office : SV 3512
Phone : +41 21 69 31751
Mail : [email protected]
Head of Unit : Priscilla Turelli
Office : AI 3151
Phone : +41 21 69 31811
Mail : [email protected]
Administrative Assistant : Séverine Reynard
Office : AI 3147
Phone : +41 21 69 31761
Mail : [email protected]